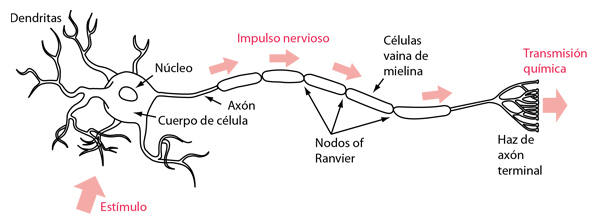
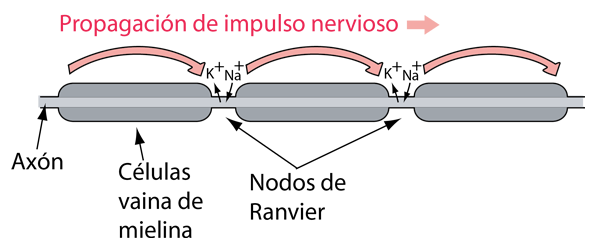
Estimulación Neural de una Fibra Muscular
Las fibras musculares se contraen por la acción de la actina y la miosina deslizándose unas sobre otras. La seńal para iniciar la contracción proviene del cerebro como parte del sistema nervioso somático..
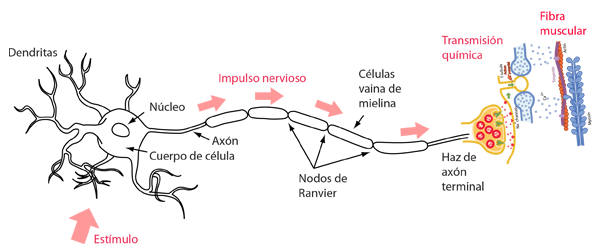
La siguiente ilustración es una representación esquemática del proceso desde la llegada de una seńal nerviosa al haz terminal del axón nervioso hasta la contracción de una fibra muscular. La estimulación de la acción muscular está asociada con el neurotransmisor químico acetilcolina.
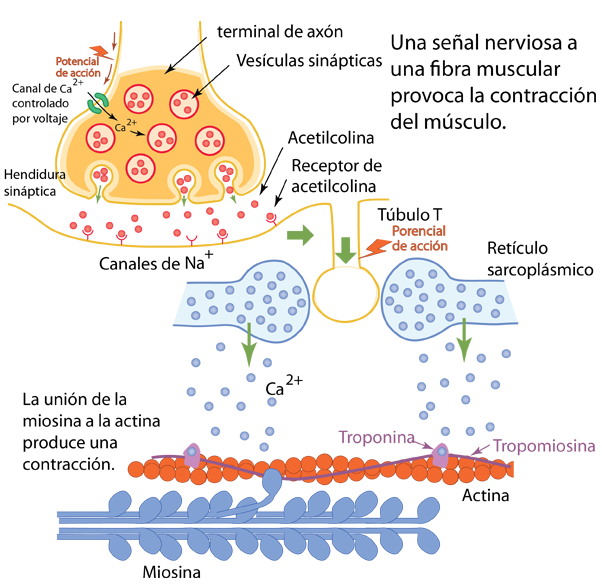
Cuando la seńal nerviosa del sistema nervioso somático llega a la célula muscular, las compuertas de calcio dependientes del voltaje se abren para permitir que el calcio ingrese a la terminal del axón. Este calcio mueve a las micelas que contienen acetilcolina para fusionarse con la membrana presináptica y liberar su acetilcolina en la sinapsis, donde se une a los receptores de acetilcolina en la superficie postsináptica. Los receptores de acetilcolina son ejemplos de receptores ionotrópicos: al unirse a la molécula de acetilcolina, abren un canal para que los iones de sodio y potasio entren en la célula. En este caso, la acetilcolina es el "elemento" que abre la puerta al sodio.
Cuando la apertura de los canales de Na envía una ráfaga de Na al interior de la célula, que, si es lo suficientemente fuerte, hace que se abran los canales de Na dependientes de voltaje cercanos y produce un potencial de acción. Este potencial de acción no se encuentra en una célula nerviosa, sino en la célula muscular.
La estructura de la fibra muscular tiene muchos tubos llamados túbulos T o túbulos transversales. Cuando el potencial de acción viaja por estos túbulos, finalmente activa las proteínas sensibles al voltaje que están vinculadas a los canales de calcio en la estructura llamada retículo sarcoplásmico (Wiki) que rodea las fibras nerviosas. Esta estructura rodeada de membrana tiene similitudes con el retículo endoplásmico de otras células. En el estado de reposo, el retículo sarcoplásmico tendrá un suministro reservado de calcio porque sus paredes tienen muchas bombas de Ca que usan energía ATP para almacenar calcio. Con el estímulo del potencial de acción, el calcio entra rápidamente en la célula e interactúa con la actina. Asociados con la actina están el complejo de troponina y la hebra de tropomiosina que bloquean la unión de la miosina. Los iones de calcio suministrados se unen a la troponina y alejan la cadena de troponina y tropomiosina "protectoras" del sitio donde puede unirse la miosina.
Para unirse a la actina, la miocina debe tener un suministro de energía, que obtiene del ATP . Habiendo absorbido energía del ATP, una unidad de la fibra de miosina estará en un estado estresado o de alta energía, como un muelle estirado. Con la acción del calcio para retirar la troponina y la tropomiosina, la estructura de la miosina puede unirse y usar la energía para tirar de la fibra de actina, acortando o contrayendo la fibra muscular.
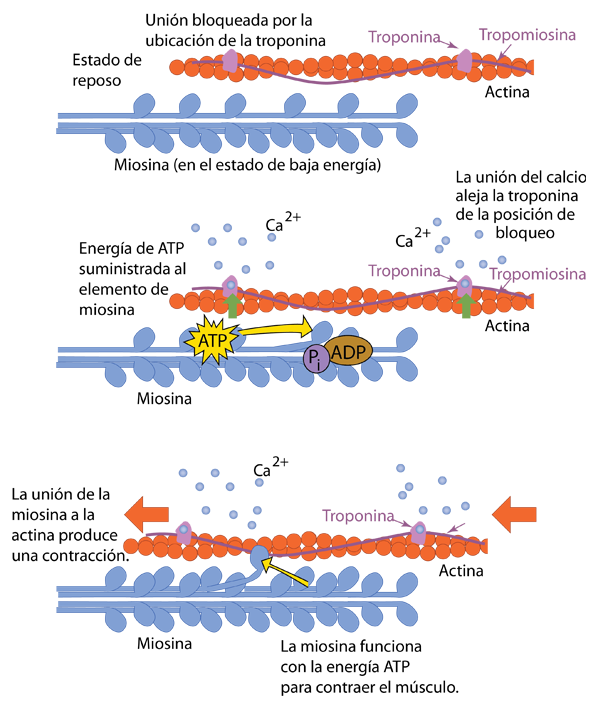
Si bien la contracción de un músculo se puede repetir siguiendo los pasos anteriores, debe haber un camino de regreso al estado de reposo, ya que no desea que sus músculos estén en un estado de contracción permanente. Se proporcionan esos mecanismos para volver al descanso. El estímulo inicial del nervio motor que inició el proceso está bajo control consciente, por lo que puede decidir relajar el músculo. La acetilcolina libre en el espacio sináptico es eliminada por otra molécula, la acetilcolinesterasa. Esta es una función esencial en las uniones neuromusculares, ya que la presencia continua de acetilcolina podría mantener el músculo en una contracción prolongada. De hecho, el papel de los venenos nerviosos y el veneno de algunas serpientes es bloquear la acción de la acetilcolinesterasa y forzar a los músculos a una estado contraído continuo. En funcionamiento normal, la acetilcolinesterasa actúa brevemente y luego cesa.
Las bombas de calcio en el retículo sarcoplásmico trabajan para recuperar el calcio y, al eliminar el calcio de los receptores del músculo, la troponina "guardaespalda" y la tropomiocina vuelven a sus posiciones de bloqueo. Las fibras de miosina y actina vuelven a su estado relajado.
|
Índice
Bioelectricidad
Tuzynski & Dixon
Sec 20.2
Neural Stimulation, lumenlearning
Frontera & Ochala
Acetylcholine receptor, Britannica
BioNinja
|